Atmung (5. Klasse, Fragen und Antworten) |
|
|
|
Home>Atmung
|
|
[Ueber diese Site][Die Tiere][Wir ueber uns][Atmung][Album-Spinnen][Album-Kaefer][Album-Wanzen][Album-Zweiflügler][Album-Hautfluegler][Album-Mikrokosmos][Album-Libellen][Tagesprotokolle]
|
|
1: Was ist innere, was äußere Atmung? Äußere A.= Transport der Atemgase Innere A (Zell-A.) = Oxidation der Energieträger zum Zweck des Energiegewinns (v.a. ATP-Bildung) 2 a.: Was ist Diffusion? 2b.: Unter welchen Voraussetzungen erfolgt der O2 Transport durch D.? 2a.: Diffusion = Verteilung von Molekülen in Gasen oder Flüssigkeiten aufgrund ihrer thermischen Eigenbewegung. Dabei gilt die Beziehung Zurückgelegter Weg ist proportional dem Quadrat der dafür benötigten Zeit. (Doppelter Weg benötigt vierfache Zeit, hundertfacher Weg benötigt zehntausendfache Zeit . . . ) Die Diff. bewirkt primär eine ständige Durchmischung der Moleküle. Sind im Ausgangszustand alle Molekültypen gleichmäßig verteilt, merkt man von der ständig ablaufenden Diff. nichts. 2b: -- ausreichend kurze Diff.-Strecke (max 1/100 mm) -- Partialdruckdifferenz (= Teildruck-Differenz, Konzentrationsunterschied). Der Teildruck eines Gases in einem Gasgemisch wird meist abgekürzt mit "P" (pressure, englisch für Druck) und Angabe der Gas-Art. "PN2" ist demnach der Stickstoff-Teildruck. -- Durchlässigkeit der zurückzulegenden Strecken für das Molekül 3: Was veranlasst den O2, von der Lungenluft ins Blut überzutreten? Der höhere P O2 in der Lungenluft gegenüber dem Blut in der Lunge 4: Wie können O2-Gehalte angegeben werden? -- Moleküle pro Liter -- Gramm pro Liter -- (Teil) Druck 5: Wovon hängt die Gasmenge, die durch Diffusion transportiert wird,
ab? Welche dieser Größen sind im
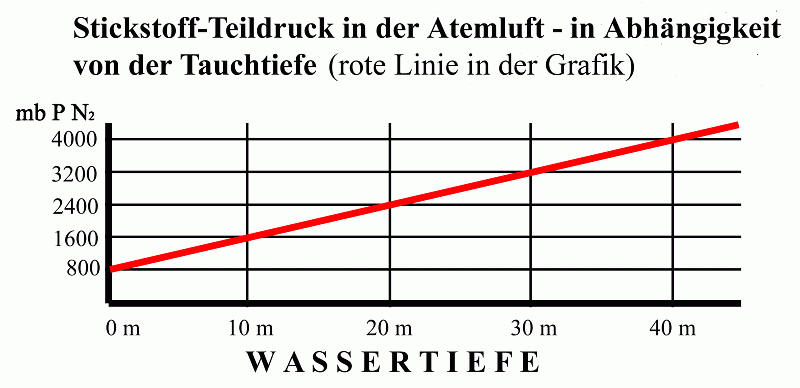
Organismus variabel, welche konstant? -- Diffusionsstrecke (konstant) -- Teildruckdifferenz (variabel) 6: Wovon hängt die Gasmenge, die in einer Flüssigkeit gelöst ist, ab? -- Teildruck -- Löslichkeit 7: Wie ist der Zusammenhang zwischen Teildruck und der absoluten Menge eines bestimmten Gases (z.B. O2) a. in der Gasphase b. in einer Flüssigkeit, wenn das Gas physikalisch gelöst ist? c. in einer Flüssigkeit, wenn das Gas chemisch gebunden transportiert wird? a: Lineare Beziehung (doppelter Teildruck = doppelte Molekülzahl) Das ist in der folgenden Grafik dargestellt am Beispiel des PN2 bei zunehmendem Luftdruck, etwa beim Tauchen
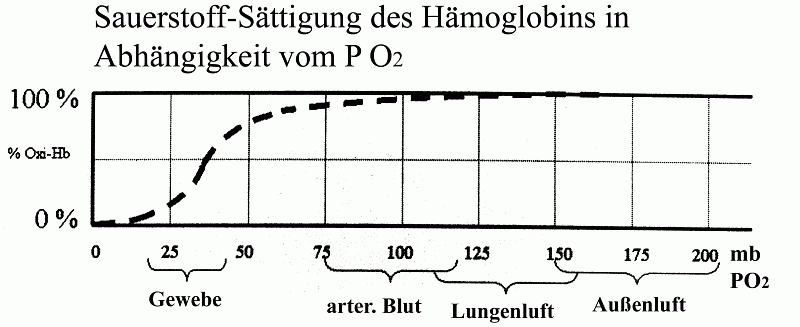
b: Vom Teildruck in der Gasphase (Lineare Beziehung: doppelter Teildruck = doppelte Zahl an gelösten Molekülen) und der Löslichkeit (doppelt so gute Löslichkeit = doppelte Molekülzahl bei gleichem Teildruck). Der Teildruck eines in einer Flüssigkeit gelösten Gases ist eigentlich ein Gleichgewichts Druck Es diffundieren ständig eine gewisse Anzahl von Molekülen von der Gasphase in die Flüssigkeit, und zwar umso mehr, je höher der Teildruck in der Gasphase ist. Gleichzeitig diffundieren ein bestimmte Menge der im Wasser gelösten O2-Moleküle in die darüberliegende Luft, und zwar umso mehr, je mehr gelöst sind und je schlechter die Löslichkeit des Gases in der Flüssigkeit ist. Ist die Zahl der ein- und ausdiffundierenden Moleküle gleich, (d.h.: es findet keine Netto-Transport mehr statt), sagen wir: Der P O2 in der Flüssigkeit entspricht dem P O2 in der darüberliegenden Luft Sprachlich ungenau, aber gebräuchlich ist die Aussage Der P O2 im Wasser beträgt xxx mb Tatsächlich wird nur ein Diffusionsgleichgewicht oder gefälle ausgedrückt und kein echter Druck innerhalb der Flüssigkeit. Die Molekülzahl O2 pro Liter Wasser kann ebenfalls eine wesentlich andere sein als die pro Liter Luft beim entsprechenden (gleichen) Druck c: Abhängig sowohl von der Affinität der Transport-Moleküle zum zu transportierenden Gasmolekül als auch vom Teildruck. Eine Besonderheit dieser Beziehung ist, dass die Affinität nicht konstant sein muss, sondern sich mit dem Teildruck ändern kann. Das bewirkt eine nicht-lineare (z.B. sigmoidale) Beziehung zwischen Teildruck und gelöster Gasmenge. Beispiel ist das Hb (Hämoglobin, roter Blutfarbstoff und Transportmolekül für Sauerstoff): Bis ca 20 mb zeigt das HB wenig Interesse am O2. Bis 50 mb nimmt dass die Beladung mit O2 stark zu und bei 100 mb sind dann bereits fast alle Hb-Moleküle mit Sauerstoff beladen. Dadurch bewirkt eine weiter Erhöhung des O2 (fast) keine Steigerung des O2-Gehaltes im Blut mehr. Die exakten Verhältnisse sind aus der Sättigungskurve abzulesen ( nachfolgende Grafik.)
Aus obiger Grafik ist gut abzulesen, dass das Hb bei dem normalerweise in der Lunge vorherrschenden PO2 praktisch zu 100 % mit O2 gesättigt ist, die stärkste Abnahme erfolgt bei den im Gewebe vorherrschenden Druckverhältnissen. Das ist kein Zufall, sondern unser Hb ist "maßgeschneidert" für die realen Druckverhältnise. Es soll voll beladen die Lunge verlassen, im Gewebe aber möglichst viel O2 abgeben. 8: Verfolge ein O2-Molekül auf dem Weg von der Luft bis zur Zelle. Wo wird es aktiv transportiert, wo durch Diff.? Außenluft ® Bronchien ® Lungenbläschen (aktiver Transport) ® Lungenbläschen ® Lungenkapillaren (Diffusion; Durchtritt durch eine 2-lagige Zellschicht: Lungenbläschen-Wand und Kapillar-Wand) ® Blutstrom (Lungenkapillaren - Lungenvene - linke Vorkammer - linke Herzkammer - Körperarterien - Körperkapillaren; aktiver Transport) - Körperkapillaren ® Zellen (Diffusion) 9: Welche O2-Teildrucke herrschen ca. in den jeweiligen Abschnitten des Transportweges der Atemgase? (Tieflage, Ruhe)
Obige Grafik zeigt nicht nur die ungefähren PO2-Verhältnisse in den jeweiligen Abschnitten des Körpers (punktierte Linien), sondern soll modellhaft auch die Engstellen und die Aufnahmekapazitäten darstellen. Dafür soll man sich das Ganze wie ein zusammenhängendes Gefäß vorstellen. Im Blut bewirkt die spezifische Sättigungskurve des Hb, dass oberhalb von 150 mb PO2 praktisch kein O2 mehr aufgenommen werden kann. 10: Welche CO2-Teildrucke herrschen ca. in den jeweiligen Abschnitten des Transportweges der Atemgase? (Tieflage, Ruhe)
Obige Grafik zeigt nicht nur die ungefähren P CO2-Verhältnisse in den jeweiligen Abschnitten des Körpers (punktierte Linien), sondern soll modellhaft auch die Engstellen und die Aufnahmekapazitäten darstellen. Dafür soll man sich das Ganze wie ein zusammenhängendes Gefäß vorstellen. Im Blut ist CO2 v.a. physikalisch gelöst, die Löslichkeit ist sehr gut. Das bewirkt, dass das "Gefäß" zylindrisch ist und einen sehr großen Durchmesser (= eine große Aufnahmefähigkeit) hat. (vgl. Fragen 6 und 7b!, beachte Unterschied zum PO2 im Blut!!! Wichtig zum Verständnis der Verhältnisse bei Hyperventilation, Frage 25) 11: Wieviel Moleküle CO2 entstehen bei der Veratmung von 1 Molekül O2? Bei Zucker genau 1 O2 = 1 CO2, bei Fett 1 O2 ca. = 0,8 CO2 12: Wieso sind die Teildruckgrößen von O2 und CO2 im Blut so verschieden? Die Vorgabe für den Teildruck eines Atemgases im Blut ist in erster Linie sein Teildruck in der Außenluft. Dieser beträgt beiO2 ca. 200 mb, bei CO2 fast 0. Außerdem ist die Löslichkeit und die Art des Transportes verschieden (siehe dazu auch 7 und 13) 13: Wieso haben wir normalerweise im arteriellen Blut noch eine so große absolute CO2 - Menge? (Sie entspricht der Menge, die bei der Veratmung der 3-fachen im arteriellen Blut vorhandenen O2-Menge entsteht) Es ist in der Lunge (fast) die gleiche CO2-Menge abzugeben, wie O2 aufgenommen wird. Da die Diffusionsstrecken für beide Gase gleich sind, hängt die Durchtrittsmenge in erster Linie vom Teildruckgefälle ab. Es baut sich demnach von selbst ein Teildruckgefälle zwischen Blut - CO2 und Außenluft - CO2 auf, das ähnlich groß ist wie bei O2. (Die bessere Löslichkeit des CO2 in Wasser bedingt eine höhere Diffusionsgeschwindigkeit des CO2 als des O2, wodurch der P CO2 sogar etwas niedriger ist als der P O2). Die viel bessere Löslichkeit des CO2 im Blut verglichen mit O2 bedingt die viel größere absolute CO2 - Menge im Blut, verglichen mit der absoluten O2 - Menge. (Vgl. 7) 14: Welche Meßgröße steuert (vorwiegend) unsere Atmung? Der pH-Wert des Blutes. Dieser wird von vielen Faktoren beeinflusst. Der Wichtigste, der sich kurzfristig verändert, ist der CO2-Gehalt. Bei kurzen intensiven Leistungen außerdem die Milchsäure (Laktat), die sich in der Muskulatur bildet. Der Sauerstoffbedarf wird also eigentlich indirekt ermittelt durch Messung des Säuregrades im Blut. Das ist bei Säugetieren (und somit auch beim Menschen, der von seiner biologischen Verwandtschaft ein Säugetier ist) sinnvoll. Denn die Endprodukte bei beiden möglichen Wegen der Energiegewinnung (Atmung: CO2, Glykolyse: Milchsäure) sind sauer. Das führt aber unter Bedingugnen, die für Menschen eigentlich unnatürlich sind, zu einigen Problemen. Zusammengefasst gilt für den Normalbetrieb also die vereinfachte Kurzformel: Die Atmung wird vorwiegend vom CO2 - Gehalt des Blutes gesteuert. Dass es auch anders geht, zeigt die Steuerung der Atmung bei Vögeln. Siehe dazu Frage 26. 15: Was ist die Restluft der Lunge? Das Luftvolumen, das nicht ausgeatmet werden kann (v.a. der Inhalt der Bronchien und der Luftröhre). Es beträgt beim Erwachsenen durchschnittlich 1 Liter. 16: Was ist das Barotrauma? Wie ist es zu erklären? Wovon hängt die Tiefengrenze, in der es eintritt, ab? Barotrauma bedeutet allgemein: Druckverletzung, also eine Verletzung an gasgefüllten Körperhohlräumen durch extremen Außendruck, z.B. durch Tieftauchen. Das Barotrauma der Lunge sei näher beschrieben: Beim Abtauchen muss sich der Gasdruck in der Lunge dem hydrostatischen Außendruck angleichen, dadurch sinkt beim Tauchen ohne Gerät das Gasvolumen in der Lunge (es kann ja keine Luft nachgeatmet werden.) Bsp: Außerhalb des Wassers: Außendruck 1 bar, Druck in der Lunge 1 bar. Einatemvolumen 5 Liter, Restluft 1 Liter = 6 Liter Gesamtvolumen. 10 m Tiefe: Außen- wie Innendruck 2 bar, Gesamtluftvolumen in der Lunge 3 Liter (2 + 1) 20 m Tiefe: Druck 3 bar, Gesamtvolumen 2 Liter (1 + 1) 50 m Tiefe: Druck 6 bar, Gesamtvolumen 1 Liter (0 + 1). Die Lunge befindet sich jetzt in maximaler Ausatemstellung!! Wird jetzt weiter abgetaucht, wird das Restluftvolumen weiter zusammengedrückt, was aber nicht ohne Verletzung abgeht (Brechen der Stützknorpel der Luftröhre, Gewebszerreißungen mit Austritt von Blut oder Lymphe in die Restluft). Die Tiefe, in der das eintritt, hängt sowohl vom Volumen der Restluft als auch vom maximal einatembaren Luftvolumen ab. (Rechenbeispiel muss durchgeführt werden können) Hinweis: Die meisten Menschen erreichen die Tiefe, bei der das Barotrauma der Lunge eintreten würde, gar nicht, da ihnen vorher der Sauerstoff ausgeht, oder sie ein Barotrauma im Bereich der Ohren oder Nebenhöhlen erleiden. 17: Wie ändert sich der Luftdruck und das Luftvolumen in der Lunge beim Abtauchen? Gibt es hierbei einen Unterschied zwischen Freitauchen und Tauchen mit Pressluft? Beim Tauchen mit Pressluft gilt natürlich ebenfalls Außendruck = Innendruck, aber das Lungenvolumen verkleinert sich nicht beim Abtauchen, da beliebig viel Luftvolumen nachgeatmet werden kann. Ein Barotrauma der Lunge kann es daher bei Tauchen mit Pressluft im Normalfall nicht geben (ausgenommen vielleicht falsch durchgeführte Notaufstiege, vgl. ausserdem 18 und 20) 18: Wodurch entsteht der Tiefenrausch? Wann bzw. wo kann er auftreten? Tiefenrausch: ist die Folge eines zu hohen P N2 im Blut (wirkt wie Narkosemittel). N2 löst sich physikalisch im Blut (und Wasser), wobei die Absolute Menge aufgrund relativ geringer Löslichkeit bei Normaldruck von 1 bar recht gering ist. Bei dem erhöhten Druck, der beim Tieftauchen gegeben ist, können aber durchaus beachtliche Mengen an gelöstem N2 erreicht werden. Da das Hineinlösen des N2 ins Blut eine gewisse Zeit dauert, ist das Auftreten des Tiefenrausches nicht nur eine Frage der Tauchtiefe, sondern auch der Tauchzeit. (Grafik). Ab 30 m muss jeder Taucher mit dem Tiefenrausch rechnen. 19: Warum können Fische in ihren natürlichen Gewässern keinen Tiefenrausch bekommen? selber überlegen . . . 20: Wodurch entsteht die Taucherkrankheit? Wann bzw. wo kann sie auftreten? Taucherkrankheit (Caisson-Krankheit) ist die Folge der Verstopfung von Kapillaren durch ausperlenden Stickstoff. Dazu kommt es, wenn -- größeren N2-Mengen sich ins Blut hineinlösen als Folge von längerem Tieftauchen -- dann schnell aufgetaucht wird, wodurch im Blut eine Übersättigung mit N2 eintritt, was eben auch zum Ausperlen von gasförmigem N2 führen kann (eine entsprechenden Effekt gibt es beim Öffnen einer Flasche mit einem kohlensäurehältigen Getränk) Die Taucherkrankheit kann tödlich enden, bei nicht-tödlichem Verlauf können lokale Gewebeschäden zurückbleiben. Sie kann auch noch viele Stunden nach dem Auftauchen eintreten. 21: Ist wettkampfmäßiges Streckentauchen ohne vorherige Hyperventilation gefährlich? Begründe! (Mit Einschränkung) nein. Denn der durch den CO2-Anstieg ausgelöste Atemreiz wird lange vor Erreichen eines kritischen O2-Mangels so stark, dass ein normal motivierter Mensch auftaucht, um zu atmen. 22: Ist wettkampfmäßiges Streckentauchen nach Hyperventilation gefährlich? Begründe Ja! Der Atemreiz wird ja hauptsächlich durch den CO2-Gehalt gesteuert. Die Hyperventilation hat aber so viel CO2 aus dem Blut entfernt, dass u.U. lange vor Einsetzen eines starken Atemreizes der O2 ausgeht und Bewusstlosigkeit eintritt (Vgl. 23) 23: Wie ändern sich die absolute und die relative (d.h. Teildruck) Menge der Atemgase im Blut bei Hyperventilation? (Getrennte Betrachtung des arteriellen und venösen Blutes sinnvoll) Hyperventilation (= absichtliches stärkeres Atmen als notwendig) führt zur Annäherung der Atemgas-Teildrucke in der Lunge an die Verhältnisse in der Außenluft (die Lunge enthält dann unnatürlich frische Luft). Für O2 heißt das: Der PO2 der Lunge steigt. Für CO2 heiß das: der P CO2 sinkt. Arterielle Sauerstoff-Situation bei Hyperventilation: Der arterielle P O2 steigt um den gleichen Betrag, wie der P O2 in der Lunge gestiegen ist. Die absolute Zahl von O2-Molekülen im Blut erhöht sich dabei aber fast gar nicht!!! Denn schon bei Normalatmung verlassen fast alle Hb-Moleküle die Lunge in O2-beladener Form. Es gibt also gar keine Aufnahmekapazität seitens des Hb mehr, sodass die Hyperventilation kein wesentlich sauerstoffreicheres Arterien-Blut erzeugen kann. Nur der in der Blutflüssigkeit physikalisch gelöste O2 - Anteil steigt proportional an, dieser Anteil ist aber insgesamt sehr gering (etwa 1/70 des an Hb gebundenen O2). Bei einem durch Hyperventilation erzeugten Anstieg des Blut - P O2 um 10 mb (z.B. von 100 auf 110 mb) werden nur sehr wenige zusätzliche O2 Moleküle ins Blut eingebracht. Arterielle CO2-Situation bei Hyperventilation: das arterielle Blut verlässt die Lunge mit einer CO2 - Beladung, die ca. 40 mb entspricht. Aufgrund der sehr guten Löslichkeit des CO2 ist die absolute Menge der CO2-Moleküle bei diesem Druck aber schon sehr groß, nämlich rund 3 mal so viel wie Sauerstoff-Moleküle im voll O2-beladenen Blut. Durch den physikalisch gelösten (und nicht chemisch gebundenen) Transport ergibt sich als weiterer Unterschied zu den O2-Verhältnissen eine streng lineares Verhältnis zwischen Blut- CO2-Menge und Teildruck. Die Hyperventilation senkt nun den P CO2 (in unserem Beispiel) um 10 mb. Dabei verlässt aber eine absolut riesige Menge an CO²-Molekülen das Blut! Bei einer durch Hyperventilation erzeugten Verminderung des Blut - P CO2 um 10 mb (z.B. von 40 auf 30 mb) werden sehr viele CO2 Moleküle aus dem Blut entfernt.
Venöse O2-Situation nach Hyperventilation: Da das arterielle Blut nicht wirklich O2-reicher war, ist das Venenblut auch bei H. O2-arm und müsste wieder neu beladen werden. Venöse CO2-Situation nach Hyperventilation: Durch die H. sind so viele CO2-Moleküle verloren gegangen, dass die CO2-Abgabe aus den Zellen ins Blut nicht ausreicht, bei einem Blutdurchgang wieder normale CO2-Verhältnise herzustellen. 25: Warum wird einem bei Hyperventilation schwindlig? Hyperventilation macht schwindlig, das ist allgemein bekannt. Weniger bekannt und durchaus verblüffend ist die Ursache. Die Erklärung ist gar nicht so einfach und setzt voraus, dass man die zugehörigen Regulationsmechanismen des Körpers kennt: Der Sauerstoffbedarf wird v.a. über die Messung des CO2-Gehaltes des Blutes festgestellt. Der Körper errechnet also den O2-Gehalt bzw. -Bedarf nach einer feststehenden Formel aus dem Ergebnis der CO2-Messung (genauer: der pH-Wert-Messung, näheres unter Frage 14) Normalerweise funktioniert das sehr gut, weil im Normalbetrieb diese beiden Größen in enger wechselseitiger Abhängigkeit stehen, so dass bei Kenntnis der einen auch die andere bekannt ist. Und doch ist dieses System störanfällig. Wird der Normalbetrieb verlassen, driften die beiden Größen auseinander. Der Körper misst aber nach wie vor den CO2-Gehalt und meint den O2-Gehalt, rechnet diesen nach dem üblichen Schema aus und gelangt prompt zu einem falschen Ergebnis! Das Gehirn ist das bezüglich O2-Mangel empfindlichste Organ. Daher gibt es eine besondere Sicherheitsmaßnahme, um bei O2-Mangel wenigstens die Versorgung des Gehirns in vollem Umfang sicherzustellen: steigt der CO2-Gehalt im Blut (was für den Körper gleichbedeutend ist mit es besteht O2-Mangel) erweitert er die Gehirn-Blutgefäße, um mehr Blut und damit mehr Sauerstoff durchs Gehirn zu leiten. Dadurch kann meistens die ausreichende Versorgung gesichert werden. Leider läuft diese Reaktion aber auch in die andere Richtung: Bei einem Zu Viel an Sauerstoff, das der Körper bei einer hyperventilations-bedingten CO2-Abnahme zu erkennen glaubt (was aber - siehe 23 - nicht der Realität entspricht), verengt er die Gehirn-Gefäße unter das Normal-Maß. Da aber der Sauerstoff-Antransport nicht besser geworden ist (siehe 23!), kommt es zu echtem Sauerstoff-Mangel im Gehirn. Dadurch und nicht durch zu viel Sauerstoff - wird man schwindlig!! Der echte Sauerstoff-Mangel im Gehirn wird aber nicht ausreichend erkannt, weil die Leitgröße der CO2-Gehalt ist ! 24. Wie ändern sich die absolute und die relative (d.h. Teildruck) Menge der Atemgase im Blut bei Aufenthalt in großer Höhe? (Getrennte Betrachtung des arteriellen und venösen Blutes sinnvoll) + 26: Welche Probleme bekommt der Körper beim Aufenthalt in großen Höhen? (über 5500 m) Mit zunehmender Höhe sinkt der Luftdruck und damit auch der O2-Teildruck. Dadurch verschlechtert sich die Sauerstoff-Sättigung des Blutes zunehmend. Die Bedingungen für die CO2-Abgabe bleiben aber immer gleich gut. Das ergibt eine Scherensituation: Atmet man bezüglich CO2 richtig, kommt es zu zunehmender O2-Unterversorgung. Verstärkt man die Atmung, um die O2-Versorgung sicherzustellen, ist das gleichbedeutend mit einer Hyperventilation bezüglich CO2. Es ist nun notwendig, 3 Höhenbereiche getrennt zu betrachten: Höhen bis ca. 1500 m Seehöhe: Keinerlei Probleme. Aufgrund der Hb-Sättigungskurve ist eine volle O2-Beladung bei Normal-Atmung sichergestellt. Höhen zwischen 1500 und 5500 m Seehöhe: Aufgrund der gleichbleibend guten CO2-Abgabebedingungen bei verringerter O2-Aufnahme bedeutet das: Atmet man "normal" (also in Bezug auf CO2 "richtig"), führt dies zu echtem O2-Mangel. Subjektiv erlebbar wird das v.a. durch ein Schwindelgefühl oder eine Euphorisierung (Auf der Alm da gibt`s koa Sünd . . .). Atmet man verstärkt (also in Bezug auf O2 "richtig"), besteht Hyperventilation bezüglich CO2. Das Schwindelgefühl bleibt, es wechselt nur die Ursache. (siehe 25). Die zu niedrigen CO2-Werte bedeuten, dass das Blut zu alkalisch ist (zu wenig Kohlensäure). Besteht der Zustand längere Zeit (Tage oder Wochen), kann sich der Körper darauf einstellen, indem er eine gewisse Menge Kalium-Ionen (Alkali-Reserve) ausscheidet. Damit wird das Blut saurer, der durch Hyperventilation bedingte Verlust an Kohlensäure wird kompensiert und das Verhältnis zwischen Atemantrieb und O2-Versorgung stimmt wieder. Das Schwindelgefühl verschwindet. (Höhenanpassung) Höhen über 5500 m Seehöhe: Die für eine volle Anpassung nötige Kalium-Abgabe führt zu Problemen anderer Art: Kalium ist nicht nur zur Regulation des Blut-pH-Wertes nötig, sondern hat viele andere Aufgaben, so ist es z.B. unerlässlich für die Nervenfunktion. Ungeachtet dessen versucht der Körper aber, soviel Kalium (unvermeidlicherweise mit Hilfe großer Wassermengen) auszuscheiden, bis seine Atemfunktion wieder im Lot ist. Verhindert man die starken Kalium-Verluste nicht durch ständige Zufuhr ausreichender Kalium- und Wasser-Mengen (bis 7 Liter täglich), kommt es zu lebensbedrohlichem Wasser- und Kalium-Mangel. Durch Zufuhr lässt sich dies verhindern. Das Bemühen des Körpers, das Kalium auf ein durch die Atemsituation bedingtes niedriges Niveau zu bringen, bleibt aber unverändert bestehen. D.h.: er scheidet unter Aufbietung seiner ganzen Energie bis 7 Liter Kalium-angereicherten Harn aus. Dies führt allmählich zu allgemeiner Erschöpfung, echte Erholung ist nicht mehr möglich. In solchen Höhen kann man sich also nur mehr aussuchen, ob man (schnell) an Wasser- und Kalium-Mangel oder (langsamer) an allgemeiner Erschöpfung stirbt. Auf jeden Fall ist die Aufenthaltszeit auf 1 2 Wochen begrenzt (Ausweg: reinen Sauerstoff atmen) Vergleich Säugetiere - Vögel beim Aufenthalt in großen Höhen Interessant ist, wenn man die Situation mit jener von Vögeln vergleicht. Insbesondere Zugvögel bewegen sich oft in sehr großer Höhe, berühmt geworden sind hierbei verschiedene Arten (z.B. Streifengänse), die regelmäßig den Himalaya in 9000 m Höhe überfliegen. Sie haben dabei offensichtlich nicht die geringsten Probleme. Wie ist das möglich? Entscheidend ist, dass Vögel den O2 Bedarf nicht über den Blut-pH-Wert ermitteln, sondern direkt messen. Dadurch kommen sie auch in der großen Höhe nicht in Verlegenheit, Messergebnisse falsch zu interpretieren. Zwar erleiden sie auch einen sehr starken CO2-Abfall, er wird aber nicht ausgewertet und spielt somit für die gesamte Atemsteuerung keine Rolle. Warum Vögel anders messen als wir mag mit der Notwendigkeit zusammenhängen, öfter innerhalb kurzer Zeitz auf große Höhen zu steigen. Wahrscheinlich ist die der wahre Hinterrund aber ein anderer: Vögel beziehen ihre Muskelleistung ausschließlich über Veratmung der Energieträger und nicht über Glykolyse. Daher fällt niemals Milchsäure an. Damit fällt aber der wichtigste Grund weg, den Blut-pH-Wert als Steuergröße für die Atmung heranzuziehen. Dass Vögel (mit ganz wenigen Ausnehmen) nicht zur Glykolyse fähig sind, mag auf den ersten Blick als Mangel erscheinen. Tatsächlich ist das aber äußerst sinnvoll: Glykolyse hat den Vorteil, dass kurzfristig sehr hohe Leistungen erbracht werden können. Der Nachteil ist aber, dass die so arbeitenden Muskeln durch die Milkchsäure schnell ermüden, was eine Erholungsphase (Pause) nötig macht. Ein Vogel kann sich aber keine Muskulatur leisten, die immer wieder eine Pause braucht. Denn das würde beim Fliegen den Absturz bedeuten. Der Verzicht auf Glykolyse bedeutet zwar den Verzicht auf kurze Höchstleistungen, aber gleichzeitig die Sicherheit, ohne Ermüdung lange durchzuhalten. (Aus dem gleichen Grund ist unser Herzmuskel nicht zur Glykolyse fähig!)
|
|
|